UNA ESPECIE DISCUTIDA.
Además de los
ejemplares clasificados en los grados erectus
o sapiens, pertenecen al interglaciar Mindel-Riss (0,4-0,2 Ma) otros
fósiles caracterizados por una capacidad craneal superior a erectus, con una caja más alta,
expansión de la región parietal y reducción del prognatismo. Estos fósiles han
sido clasificados bajo distintos nombres:
-
Heidelbergensis.
-
Rodhesiensis
-
Erectus petraloniensis
-
Swanscombensis
-
Soloensis
-
Etc.
Las alternativas son:
1. Atribuirlos al grado erectus.
a.
La
solución parsimoniosa es colocarlos todos dentro de erectus.
b.
Para
Stringer (1993) la falta de rasgos derivados propios del erectus en los ejemplares de Atapuerca, confirma su tesis de que
nunca hubo erectus en Europa y
sugiere que pertenecen ya al taxón de los neandertales.
2. Atribuirlos a la especie Homo heidelbergensis. Para Rightmire (1997)
heidelbergensis es un taxón válido que
quizá sea el último antecesor común de neandertales y humanos modernos, con una
evolución por separado en Europa y África. Para Ward y Stringer la población europea
lleva hasta los neandertales y la africana a los sapiens.
3. Atribuir los restos africanos a rhodesiensis (luego a helmei) y los europeos a:
a. heidelbergensis. No todos los fósiles de heidelbergensis presentan rasgos neandertales
en la misma parte anatómica El
equipo de Atapuerca (2011) considera que heidelbergenesis
es un grado, un conjunto de grupos con evolución en paralelo. Los
neandertales pudieron surgir del grupo de Atapuerca o de otro.
b.
neanderthalensis. Stringer (1993) y Arsuaga consideran que la
gran variación de la muestra de Atapuerca hace difícil seguir sosteniendo el
taxón heidelbergensis porque esta
especie y los neandertales se solaparían. Como mucho heidelbergensis sería un ancestro exclusivo de los neandertales.
Los homínidos europeos del Pleistoceno Medio formarían una cronoespecie y
serían antecesores solo de los neandertales, que derivarían por anagénesis. Esto
obligaría a extender neanderthalensis a
través de un periodo muy largo, en el que hubo cambios morfológicos notables.
Dean y colaboradores (1998) definen cuatro estadios dentro de la evolución
anagenética de los neandertales.
c.
Para
Aguirre y Lumley (1977), en el Pleistoceno Medio existieron tres grupos
genética y geográficamente separados, que no pueden ser considerados erectus (que quedaría restringido a Java
y no contribuiría a la aparición de neandertales ni sapiens), aunque su cercanía supone un argumento a favor de la
hipótesis multirregional erectus-sapiens:
i. Lejano Este (Zhoukoudian). La posibilidad de
coexistencia en Java de erectus y sapiens ha sido propuesta por Swisher y
colaboradores (1996). Para ellos, los erectus
sobrevivieron en Java al menos 0,25 mda después de haberse extinguido en
China. Sugieren incluso que erectus y
sapiens pudieran intercambiar genes.
De clasificar el espécimen hallado en Jinniu Shan (0,2 mda) como sapiens, las dos especies coexistirían
también en China.
ii. Magreb.
iii. Grupo anteneandertaliense: Mauer, Arago,
Atapuerca, Montmaurin, Bañolas.
EVOLUCIÓN DE HOMO DURANTE EL INTERGLACIAR MINDEL-RISS
|
África
|
Europa
|
Asia
|
Solución parsimoniosa
|
Grado erectus
|
Grado erectus
O grado sapiens
|
Grado erectus
|
Aceptar heidelbergensis en África y Europa
|
Heidelbergensis = pre-sapiens
|
Heidelbergensis = pre-neandertales o neandertales
|
Grado erectus
|
Aceptar heidelbergensis solo en Europa
|
¿Pre-sapiens como cladogénesis de antecessor?
|
Heidelbergensis = pre-neandertales o neandertales
|
Grado erectus
|
Aceptar helmei
|
Rodhesiensis, después helmei (último antecesor común de neandertales y sapiens)
|
Heidelbergensis, después neandertales y helmei
|
Grado erectus, después helmei
|
Yacimientos
El
grado heidelbergensis abarcaría los hallazgos europeos datados entre 0,7-0,134
Ma en los que predominan rasgos primitivos, aunque muestren algunos caracteres incipientes
que indican que son los antepasados de los neandertales. Estos rasgos de neandertales
se presentan de forma frecuencial y en mosaico.
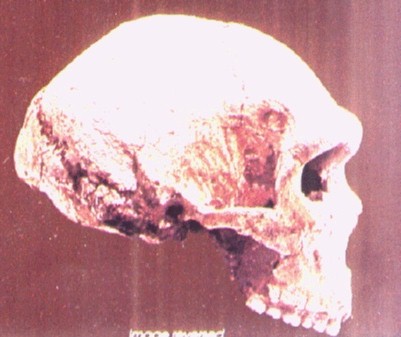
"Heidelberg
Man", "Mauer Jaw", Descubierto
por trabajadores de una cantera en 1907, cerca de Heildelberg (Alemania), su edad
se estima en 0,6 Ma. Se trata de una mandíbula inferior, con el mentón retraído
y todos los dientes. Muelas relativamente pequeñas, sobre todo M3. La mandíbula
es extremadamente larga y robusta, como la del
Homo erectus, pero con los dientes más pequeños.
Arago (Francia), yacimiento con
veinte niveles de ocupación entre 0,55-0,34 Ma, bajo la dirección de Henry y
Marie-Antoinette de Lumely. 80 fósiles. Sobresalen un cráneo y dos mandíbulas
·
Arago
21,
Tautavel Man. Descubierto en 1971
por Henry de Lumley, datado en 0,4 Ma. Rostro casi completo, con 5 dientes molares
y parte de la bóveda craneal. Su capacidad craneal es de 1.150 cc. Bóveda
craneana baja, torus supraorbitario grueso con depresión glabelar neta; huesos
malares y rostro macizos, sin fosa poscanina; el frontal es estrecho y la angulación
del occipital acusada; la sección transversal de la bóveda, ligeramente
tectiforme; las líneas temporales, altas y marcadas.
·
Arago 2. Mandíbula de un individuo
femenino.
·
Arago 13. Mandíbula de un individuo
masculino.
·
Dientes de 10 individuos distintos.
·
Arago 44. Hueso ilíaco. En conjunto con
otros huesos nos permite estimar una talla de 1,64 m.
Vergranne (Francia) canino de
menos de 0,5 mda.
Fontana Ranuccio (Italia) Dos
dientes incisivos datados en 0,458 mda.
Cava Pompi (Italia). Fragmentos
de cráneo, cúbito y tibia. 0,45 mda.
Visogliano (Italia). Mandíbula y
cinco dientes sueltos. 0,4 mda.
Vanosa-Notarchirico (Italia).
Fémur asociado a industria de modo 2. No se ve técnica de Levallois. 0,4-0,35
mda.
Bilzingsleben (Alemania).
Fragmentos de cráneo asociados a industria de modo 1 evolucionado. 0,4 mda.
Vérteszöllös (Hungría).
Occipital y varios dientes. 0,38-0,35 mda.
Cueva Negra del Estrecho del Río
Quípar (España) 0,45-0,35 mda. Fragmentos de de húmero y cúbito humano y
dientes.
Orgnac 3 (Francia). Decena de
dientes aislados, parecidos a los de Arago. Aparece técnica de Levallois.
0,35-0,32 mda.
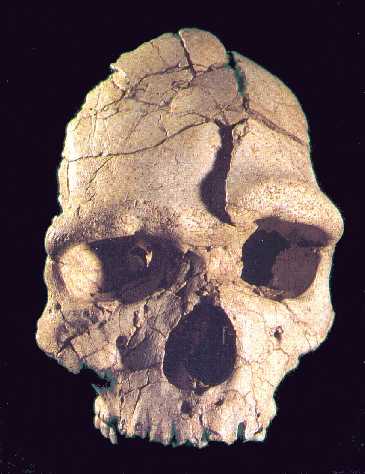
Petralona
1. Cráneo casi completo. Apomorfias neandertales en órbitas, rostro hacia
delante, frente y bóveda craneanas hacia atrás con una cara alargada y una
mandíbula muy ancha. Descubierto por habitantes de Petralona (Grecia) en 1960. Su
capacidad craneana es de 1220 c.c., alto para ser
erectus, pero bajo para
ser
sapiens. Su edad se estima entre 0,25-0,15 mda. Hay quien considera que
se trata del último
Homo erectus, y otros le ven características de neandertal.
Atapuerca. Sima de los
Huesos – Ibeas de Juarros. Más de 5.000 fósiles (90% de los conocidos del
Pleistoceno Medio en todo el mundo) de unos 28 individuos. 0,32 mda. Los
primeros, descubiertos en 1976, por el equipo de T. Torres. En su mayor parte,
corresponden a individuos jóvenes y adolescentes. El peso calculado por J. M.
Carretero para jóvenes y adultos, está entre 58,75 y 81,43 kg. La estatura
media de los varones, 1,74 m y la de las hembras 1,6 m. Para Emiliano Aguirre
(2008) estos individuos se hallaban juntos en el interior de una cueva cuya
entrada se cerró accidentalmente. Los restos fueron arrastrados por el fango y
se deslizaron hasta la boca de la sima. Por el camino, algunos carnívoros
penetraron por alguna rendija y dejaron sus huellas en los huesos. Esto explicaría
también la ausencia de niños pequeños, cuyos restos habrían sido totalmente
engullidos por los carroñeros. Los individuos, en general, estaban sanos aunque
habían pasado por algunas crisis alimentarias; eran propensos a las afecciones
artrósicas y a algo parecido a la osteoporosis; no hay rastro de accidente o
enfermedad causante de la muerte.
·
16 cráneos. 10
individuos padecían cribra orbitaria, extensión de perforaciones en el
techo de la cavidad de las órbitas. En varios casos se observan traumatismos
curados.
·
20 mandíbulas (medio
centenar de piezas), cuatro de ellas enteras. Son robustas, sin mentón. Cuerpo
mandibular más bien bajo, con el borde inferior ancho, plano, paralelo a la
fila de dientes y con un acusado reborde. Ramas ascendentes largas, poco altas
y con el borde superior poco escotado. En seis individuos se observa una
artrosis degenerativa de la articulación del hueso temporal y la mandíbula.
·
Más de 500 dientes
o Incisivos superiores en pala.
o Caninos con cínguos gruesos.
o Premolares no molariformes.
o Molares decrecientes, ya del primero al segundo, tanto en largo (igual
que los neandertales) como en ancho (típicamente moderno).
o Son frecuentes las hipoplasias en el esmalte. No se ven signos de
caries, que aparecerían con el consumo de cereales, pero sí de la utilización
de mondadientes. Hay marcas en la superficie externa de los dientes anteriores
causadas por filos silíceos al utilizar la dentadura como herramienta.
o Para María Martinón Torres y José María Bermúdez de Castro (2011) la
morfología de estos dientes es completamente neandertal y por ello estos
individuos de Atapuerca pertenecen a un grupo o especie diferenciados, pues
estos rasgos están ausentes en los restos de los demás yacimientos de similar
datación.
·
42 restos de clavículas,
114 de escápulas, 645 restos de vértebras, 466 de costillas.
o Las clavículas tienen como rasgo distintivo su longitud, su fuerte
curvatura en ese y el fuste comprimido de modo anteroposterior, con sección
ovoide.
o Conducto medular largo en los atlas y corto en los axis, que son anchos
en comparación con los modernos.
·
110 restos de húmeros,
65 de cúbitos y 85 de radios.
o En el húmero, la cabeza articular es oval con un ángulo de torsión
bajo. Varios refuerzos en la zona articular del codo.
·
173 restos de fémures,
20 de rótulas, 89 de tibias y 105 de peronés. Los huesos largos son muy
fuertes.
·
121 huesos del carpo,
103 del tarso, 93 metacarpianos, 80 metatarsianos, 208 falanges de pies y 304
falanges de manos. En los restos de niños, se dan varios casos de líneas de
Harris, consecuencia típica de enfermedades infantiles.
·
Hioides y huesecillos
del oído. I. Martínez ha deducido de su análisis la capacidad de habla de
aquellos individuos.
·
Elvis. 0,5 mda. Pelvis
muy ancha, con fuertes inserciones musculares correspondientes a una piernas
muy robustas. La pelvis mejor conservada de todo el registro fósil, junto con
otros restos del esqueleto, correspondiente a un varón muy corpulento que
padeció importantes enfermedades degenerativas desde mucho antes de morir, con
más de 45 años de edad. Dichas enfermedades debieron de producirle intensos
dolores en las zonas lumbar y pélvica que le obligaban a caminar encorvado y,
probablemente, a utilizar un bastón para mantenerse erguido. Debido a sus
mermadas condiciones físicas, este individuo no estaba en condiciones de cazar
ni de llevar a cabo otras tareas útiles para su comunidad, lo cual ha llevado a
la conclusión de que, además de alimentación, el anciano debió también de
recibir ayuda para seguir el ritmo de la marcha de su gente, un grupo de
cazadores nómadas que se desplazaban junto a las manadas de herbívoros de los
que se alimentaban. Un tipo de conducta social de ayuda a los más desvalidos de
la que existen muy pocos indicios para épocas tan remotas pero sobre la que en Atapuerca
ya se ha encontrado más de una prueba.
o En efecto, en marzo de 2009 se publicó un estudio que demostraba que un
niño con el cráneo deforme, por fusión prematura de los huesos del cráneo, de unos
10 años (que los científicos bautizaron como Benjamín) y que pertenecía
a la misma especie y época del anciano, fue cuidado y mantenido con vida todo
el tiempo posible por sus congéneres.
·
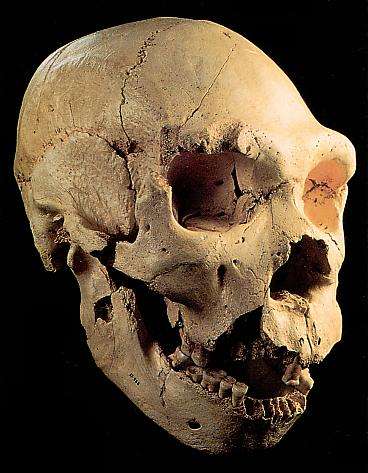
Miguelón, Atapuerca
5 (AT 700). Descubierto en la Sima de los Huesos (Atapuerca) entre 1992 y 1993 por
Juan Luis Arsuaga. Su antigüedad se estima en 0,3 Ma; su capacidad craneal es de
1.125 c.c. Se trata del cráneo más completo de heilderbengensis descubierto hasta ahora. (Arsuaga et al. 1993; Johanson
and Edgar 1996). Se ha identificado la mandíbula que le corresponde.
o Macizo
maxilar grande, proyectado hacia delante sin fosa canina entre el saliente
nasal y el pómulo ni fosa maxilomalar entre el pómulo y la pared de las raíces
de los molares superiores. Orbitas grandes y casi redondas. Prognatismo y
adelantamiento del arco de los dientes que deja un espacio retromolar entre el
talón de la última muela inferior y el borde anterior de la rama ascendente de
la mandíbula.
o Padecía
varias alteraciones en la boca. La más grave, una infección en el lado
izquierdo tras la fractura de un diente.
o Del
estudio de la lateralidad del cerebro, se concluye que era diestro.
·
Atapuerca 4. Caja craneal completa, sin
su parte facial. Se observa una artritis avanzada en los cónditos occipitales,
con importante dificultad y dolor en los movimientos de la cabeza y los dos
conductos auditivos obstruidos por hiperostosis.
·
El frontal huido, la bóveda baja y el
occipital alargado y anguloso hacia atrás del cráneo 5 no son compartidos en el
cráneo 4. La bóveda relativamente baja y el frontal estrecho son rasgos
comunes.
Atapuerca-Galería, en la
comunicación con la Sala de los Zarpazos. 0,3 Ma.
·
AT 76-T1H. Fragmento de
mandíbula hallado por Trinidad Torres (1976). La presencia de espacio
retromolar, el toro mandibular y los tamaños de los últimos molares jutifican
su asignación al demo de los preneandertales europeos.
·
Fragmento de cráneo
hallado en 1995 que comprende la parte posteromesial del parietal derecho,
parte posterior de la sutura sagital, otro fragmento menor del parietal
izquierdo y el tramo central de la sutura labdática, con una porción muy
pequeña adherida del occipital que incluye el lambda. El parietal es muy grueso
y fue hundido por un golpe que luego sanó.
Boxgrove (Reino Unido).
Excavado por Mark Roberts en los 90. El yacimiento se mantuvo intacto desde que
hace 0,5 Ma fuese cubierto de forma rápida por el fango, que permitió su
conservación. Se han encontrado huesos de elefante, rinoceronte, caballo,
ciervo, león, lobo, hiena, castor, liebre, ratón de campo y rana. Muchos huesos
conservan huellas líticas y se ha podido reconstruir el proceso de matanza o
despiece. Para Roberts, los animales fueron cazados mientras que Gamble
defiende que son fruto de la carroña. Los únicos restos humanos son dos dientes
y parte de una tibia (0,35-0,3 Ma). No hay hogares ni señales de estructuras.
Castel di Guido
(Italia). Fragmentos craneales. Una diáfasis de fémur. Asociados a industria
modo 2. 0,3 Ma.
Ponte Mammolo (Italia).
Un fémur. 0,2 Ma.
Pech-de-l’Aze (Francia).
Infantil. 0,4-0,2 Ma.
Biache-Saint Vaast
(Francia) Cráneo incompleto. Trazado de la red meníngea avanzado, parecido con
Swanscombe y con neandertal. 0,263-0,175 mda.
Swanscombe (Reino
Unido). Parietal y occipital de hembra joven adulta. 1.300 cc. Formas muy
próximas a neandertal. 0,25 mda.
Pontnewydd (Reino Unido)
Fragmentos de maxilar y tibia. Dientes. 0,251-0,195 mda.
Steinheim (Alemania)
Cráneo casi completo, deformado y restaurado. Apomorfias de neandertal poco
avanzadas. 0,225 mda.
Ehringsdorf (Alemania)
Restos craneales y poscraneales de varios individuos. Rasgos de neandertal
avanzados. 0,25-0,2 mda.
Grotta del Principe
(Italia). Pelvis de individuo femenino. 0,2 mda.
Sedia del Diavolo
(Italia). Fragmentos de cráneo y fémur y un metatarsiano. 0,2 mda.
Grotta del Poggio
(Italia). Un diente y varios poscraneales. 0,2 mda.
Casal de Pazzi.
Fragmento de parietal. 0,2 mda.
Le Lazaret (francia).
Paietal de inmaduro, con cicatriz de trauma y meningioma postraumático; un
incisivo lacteal y un canino adulto. Asociado a industria modo 2 evolucionado.
0,16 mda.
Apidima I (Grecia). Dos
cráneos. 0,2 mda.
Apidima II (Grecia).
Rostro, frente y bóveda craneal muy próximos a neandertales de Saccopastore y
Gibraltar.
Grotte Suard. La Chaise
(Francia). Pates de un cráneo adulto, otro infantil y su mandíbula, tres
dientes. Asociados a industria tipo 2 evolucionado, con lascas Levallois. 0,13
mda.
Cueva del Congosto
(Alcorlo, Guadalajara, España). Un metatarsiano y una vértebra cervical.
Asociados a restos de animales con trazas de haber sido un basurero de hienas.
Cueva de Lezetxiki
(Mondragón, Guipúzcoa, España). Húmero completo de un individuo femenino.
Datación incierta.
Reilingen (Alemania).
Cráneo incompleto. Algunos rasgos afines a neandertal.
Montmaurin (Francia).
Mandíbula. Rasgos de preneandertal.
Características.
- Dientes con tamaño y corona reducida, mucho más pequeños que el erectus.
- Caja craneal voluminosa y en pendiente. Frente deprimida, con toro
poderoso en dos partes. Bóveda craneal bastante baja y alargada. Occipucio
anguloso con repliegue.
- Prognatismo profundo. Arco cigomático saliente y orientado hacia
delante.
- Arcada dental parabólica. Mandíbula moderadamente robusta.
- Reducción del tamaño de los molares.
- Pelvis corta y en forma de cubeta. Miembros inferiores
relativamente largos.
Inferencias sobre su modo de vida.
- Regiones
templadas o cálidas. Paisajes variados de árboles o agua, pero no jungla
densa.
- Modo de vida y
dieta: cazadores capaces de matar animales grandes mediante emboscada; recolectores
de pequeños animales, huevos y todo tipo de plantas silvestres comestibles.
De su higiene sabemos que usaban palillos de dientes para la limpieza de los
espacios interdentales. Usaban abrigos de piel para evitar el frío y fabricaban
herramientas de piedra y madera. En Schöningen se hallaron lanzas de
madera datadas en 0,4 Ma.
- Campamentos: construcción
de cabañas de madera y quizás pieles de animales, pero también uso de abrigos
y vestíbulos de cuevas.
- Organización social:
pequeños grupos o clanes de entre 15 y 40 individuos jerarquizados, que explotaban
y defendían los recursos de un territorio y que realizaban contactos esporádicos
con otros grupos para evitar la endogamia. Probablemente podían
comunicarse mediante el habla.
Más sobre Homo heidelbergensis y Homo neanderthalensis.
















{kind=link}
{kind=link}
{kind=link}
No hay comentarios:
Publicar un comentario