 |
| Tony Bennet. Lady Gaga. Vanity Fair. Enero 2012. |
miércoles, 30 de noviembre de 2011
Lady Gaga desnuda por Tony Bennet.
martes, 29 de noviembre de 2011
Filogénesis del conducto vocal supralaríngeo.
La facultad de hablar está
relacionada con una determinada configuración anatómica del tramo final de la
laringe. Como indica Lieberman, la longitud del tubo formado por la boca debe
ser equivalente a la del otro tubo de la parte posterior de la lengua, ya
dentro de la faringe, para que se puedan producir los sonidos del habla humana.
Como resultado, solo una posición baja de la laringe permite vocalizar. Esto
conlleva ciertas complicaciones a la hora de respirar y tragar al mismo tiempo. Los
neonatos pueden mamar y respirar al mismo tiempo por la situación muy alta de su
laringe, que es muy similar a la de los chimpancés. La laringe “baja” en el cuello
en los humanos a los dos años aproximadamente, edad en que los niños empiezan a
articular palabras. Todos los primates tienen un saco de aire adherido a una
extensión del hioides en forma de copa (bulla), excepto los seres humanos, en
los que se ha reducido a un órgano vestigial. En el Australopithecus afarensis estaba presente esta bulla, pero ya
estaba ausente en el Homo heidelbergensis.
Según los estudios de Bart de Boer (2011), la presencia de este saco hace
dificulta la vocalización.
- Para
Aiello, la posición baja de la laringe es una consecuencia de las adaptaciones
anatómicas necesarias para la bipedación. Otra consecuencia de la bipedación es
la liberación de la laringe de las tareas locomotrices que desempeña en los
simios que la utilizan para dar firmeza a pecho y brazos. Al quedar liberada la
laringe de esas funciones, las cuerdas vocales pudieron hacerse más membranosas
y se capacitaron así para emitir una mayor variedad de sonidos.
-
Laitman (1984) sostiene que los australopitecinos
disponían de una laringe en posición alta. El descenso comenzaría en erectus.
-
Lieberman, a partir de las marcas dejadas por los
músculos, ha afirmado que el habla sería un fenómeno muy tardío, propio de los
seres humanos de aspecto moderno y, en parte, de los neandertales. Estos
últimos serían capaces de emitir parte de los sonidos al alcance de los
humanos, pero no todos
-
Krantz (1988) piensa que un descenso laríngeo completo
sólo se realizó hace 40.000 años (¡coincide con la explosión simbólico-artística
del Paleolítico!). La primera fase, hace 200.000 años, habría modernizado el
cráneo y especialmente la laringe, dando lugar a una cavidad que supondría la
mitad de la actual y permitiría una conducta vocal imperfecta.
El hallazgo en Kebara (Oriente Próximo) de un hueso
hioides fosilizado (un hueso intermedio entre la laringe y la base del cráneo que
da lugar a gran parte de la musculatura lingual y laríngea), perteneciente a un
Homo neanderthalensis, nos permite
sostener que su aparato fonador era similar al nuestro. Lieberman ha criticado
la identificación entre hueso hioides y capacidad de habla. Pero la mayor parte
de las especulaciones sobre el lenguaje de los homínidos se basan en la conducta
simbólica (enterramientos y objetos de adorno) y por tanto en el cerebro.
En la ducha por el equipo.
 |
| Campaña promocional del Bohemias 1905 de Praga. 2011. |
De nuevo un equipo deportivo utiliza cuerpos desnudos como forma de promoción. En este caso se trata del Bohemias 1905 de Praga. El propio presidente del club, Anton Doll, realizó las fotografías. Ha colaborado Panenka, directivo del club y autor del famoso penalti que dio a Checoslovaquia el campeonato de Europa de fútbol de 1976.
 |
| Panenka. Promoción del Bohemians 1905. 2011. |
 |
| Campaña promocional del Bohemias 1905 de Praga. 2011. |
 |
| Campaña promocional del Bohemias 1905 de Praga. 2011. |
lunes, 28 de noviembre de 2011
La cultura de Huajomulco.
 |
| Huajomulco. Pirámide. |
Muchas de las figuras son personas sentadas con sus manos sobre las rodillas y algunos tienen sombreros con serpientes. Las esculturas están hechas de un material que combina arena, limo y agua, y pintadas de azul o verde y se cree que son del período epiclásico, entre el año 600 y el 900.
Thomas Charlton, arqueólogo estadounidense de la universidad de Iowa, que trabaja en la zona, también cree que se trata de una nueva cultura que existió entre la caída de Teotihuacan y el comienzo del poderío Tolteca.
http://news.nationalgeographic.com/news/2008/12/081208-mexico-pyramid.html
domingo, 27 de noviembre de 2011
Miru Kim. Viviendo como los cerdos.
 |
| Miru Kim. The pig that therefore i am. |
Miru Kim nació en Stoneham, Massachusetts en 1981, pero se crió en Seúl, Corea. Es hija del filósofo Kim Young-Roble. Regresó a Massachusetts en 1995 para estudiar en la Phillips Academy y en 1999 se trasladó a Nueva York para estudiar en la Universidad de Columbia.
Durante la próxima feria de arte Art Basel Miami Beach (6 al 9 de diciembre), Miru vivirá entre los cerdos, para provocar una reflexión sobre las similitudes y diferencias entre el ser humano y una de sus víctimas culinarias preferidas.
http://www.mirukim.com/
http://jmflseixalbo.blogspot.com/2011/06/miru-kim-explorando-desnuda-las-ruinas.html
http://www.artbaselmiamibeach.com/go/id/xa/
 |
| Miru Kim. The pig that therefore i am. |
 |
| Miru Kim. The pig that therefore i am. |
http://www.mirukim.com/
http://jmflseixalbo.blogspot.com/2011/06/miru-kim-explorando-desnuda-las-ruinas.html
http://www.artbaselmiamibeach.com/go/id/xa/
sábado, 26 de noviembre de 2011
Mookie Tenembaum: Inmortalidad o muerte.
 |
| Mookie Tenembaum. Inmortalidad o Muerte. |
La intención del artista es poner en evidencia las manifestaciones y protestas urbanas como forma de organización social reclamando al Congreso algo tan absurdo como una modificación a la Constitución que garantice el derecho a la inmortalidad.
“¡No podemos morir! Estamos muy ocupados”.
 |
| Mookie Tenembaum. Inmortalidad o Muerte |
http://noticias.lainformacion.com/arte-cultura-y-espectaculos/artes-general/un-artista-argentino-lleva-a-la-calle-a-modelos-semidesnudas-con-boinas-del-che_OYhFGNnCE0J7d968VjzEZ4/
http://www.clarin.com/sociedad/Desfile-derecho-inmortalidad_0_598140351.html
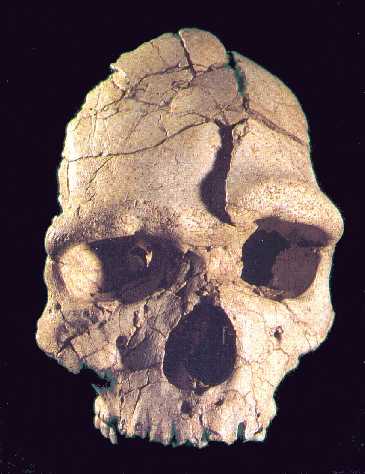
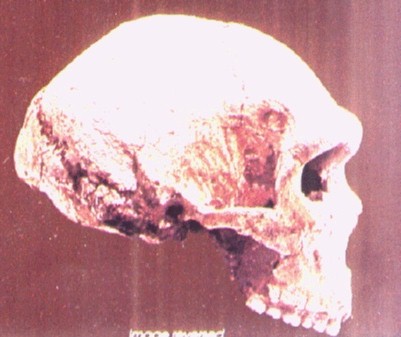
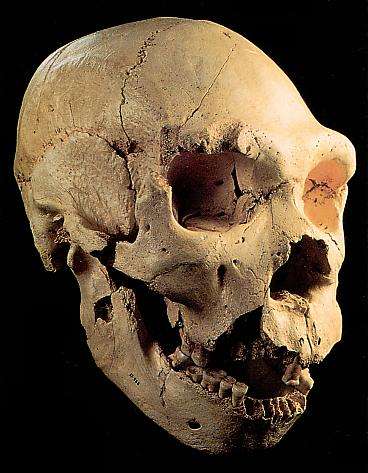
El grado Heidelbergensis. Demo de Mauer-Arago.
 |
| Homo heidelbergensis. Atapuerca 5. |
UNA ESPECIE DISCUTIDA.
Además de los ejemplares clasificados en los grados erectus o sapiens, pertenecen al interglaciar Mindel-Riss (0,4-0,2 Ma) otros fósiles caracterizados por una capacidad craneal superior a erectus, con una caja más alta, expansión de la región parietal y reducción del prognatismo. Estos fósiles han sido clasificados bajo distintos nombres:- Heidelbergensis.- Rodhesiensis- Erectus petraloniensis- Swanscombensis- Soloensis- Etc.Las alternativas son:1. Atribuirlos al grado erectus.a. La solución parsimoniosa es colocarlos todos dentro de erectus.b. Para Stringer (1993) la falta de rasgos derivados propios del erectus en los ejemplares de Atapuerca, confirma su tesis de que nunca hubo erectus en Europa y sugiere que pertenecen ya al taxón de los neandertales.2. Atribuirlos a la especie Homo heidelbergensis. Para Rightmire (1997) heidelbergensis es un taxón válido que quizá sea el último antecesor común de neandertales y humanos modernos, con una evolución por separado en Europa y África. Para Ward y Stringer la población europea lleva hasta los neandertales y la africana a los sapiens.3. Atribuir los restos africanos a rhodesiensis (luego a helmei) y los europeos a:a. heidelbergensis. No todos los fósiles de heidelbergensis presentan rasgos neandertales en la misma parte anatómica El equipo de Atapuerca (2011) considera que heidelbergenesis es un grado, un conjunto de grupos con evolución en paralelo. Los neandertales pudieron surgir del grupo de Atapuerca o de otro.b. neanderthalensis. Stringer (1993) y Arsuaga consideran que la gran variación de la muestra de Atapuerca hace difícil seguir sosteniendo el taxón heidelbergensis porque esta especie y los neandertales se solaparían. Como mucho heidelbergensis sería un ancestro exclusivo de los neandertales. Los homínidos europeos del Pleistoceno Medio formarían una cronoespecie y serían antecesores solo de los neandertales, que derivarían por anagénesis. Esto obligaría a extender neanderthalensis a través de un periodo muy largo, en el que hubo cambios morfológicos notables. Dean y colaboradores (1998) definen cuatro estadios dentro de la evolución anagenética de los neandertales.c. Para Aguirre y Lumley (1977), en el Pleistoceno Medio existieron tres grupos genética y geográficamente separados, que no pueden ser considerados erectus (que quedaría restringido a Java y no contribuiría a la aparición de neandertales ni sapiens), aunque su cercanía supone un argumento a favor de la hipótesis multirregional erectus-sapiens:i. Lejano Este (Zhoukoudian). La posibilidad de coexistencia en Java de erectus y sapiens ha sido propuesta por Swisher y colaboradores (1996). Para ellos, los erectus sobrevivieron en Java al menos 0,25 mda después de haberse extinguido en China. Sugieren incluso que erectus y sapiens pudieran intercambiar genes. De clasificar el espécimen hallado en Jinniu Shan (0,2 mda) como sapiens, las dos especies coexistirían también en China.ii. Magreb.iii. Grupo anteneandertaliense: Mauer, Arago, Atapuerca, Montmaurin, Bañolas.EVOLUCIÓN DE HOMO DURANTE EL INTERGLACIAR MINDEL-RISS
Yacimientos
El grado heidelbergensis abarcaría los hallazgos europeos datados entre 0,7-0,134 Ma en los que predominan rasgos primitivos, aunque muestren algunos caracteres incipientes que indican que son los antepasados de los neandertales. Estos rasgos de neandertales se presentan de forma frecuencial y en mosaico."Heidelberg Man", "Mauer Jaw", Descubierto por trabajadores de una cantera en 1907, cerca de Heildelberg (Alemania), su edad se estima en 0,6 Ma. Se trata de una mandíbula inferior, con el mentón retraído y todos los dientes. Muelas relativamente pequeñas, sobre todo M3. La mandíbula es extremadamente larga y robusta, como la del Homo erectus, pero con los dientes más pequeños.Arago (Francia), yacimiento con veinte niveles de ocupación entre 0,55-0,34 Ma, bajo la dirección de Henry y Marie-Antoinette de Lumely. 80 fósiles. Sobresalen un cráneo y dos mandíbulas· Arago 21, Tautavel Man. Descubierto en 1971 por Henry de Lumley, datado en 0,4 Ma. Rostro casi completo, con 5 dientes molares y parte de la bóveda craneal. Su capacidad craneal es de 1.150 cc. Bóveda craneana baja, torus supraorbitario grueso con depresión glabelar neta; huesos malares y rostro macizos, sin fosa poscanina; el frontal es estrecho y la angulación del occipital acusada; la sección transversal de la bóveda, ligeramente tectiforme; las líneas temporales, altas y marcadas.· Arago 2. Mandíbula de un individuo femenino.· Arago 13. Mandíbula de un individuo masculino.· Dientes de 10 individuos distintos.· Arago 44. Hueso ilíaco. En conjunto con otros huesos nos permite estimar una talla de 1,64 m.Vergranne (Francia) canino de menos de 0,5 mda.Fontana Ranuccio (Italia) Dos dientes incisivos datados en 0,458 mda.Cava Pompi (Italia). Fragmentos de cráneo, cúbito y tibia. 0,45 mda.Visogliano (Italia). Mandíbula y cinco dientes sueltos. 0,4 mda.Vanosa-Notarchirico (Italia). Fémur asociado a industria de modo 2. No se ve técnica de Levallois. 0,4-0,35 mda.Bilzingsleben (Alemania). Fragmentos de cráneo asociados a industria de modo 1 evolucionado. 0,4 mda.Vérteszöllös (Hungría). Occipital y varios dientes. 0,38-0,35 mda.Cueva Negra del Estrecho del Río Quípar (España) 0,45-0,35 mda. Fragmentos de de húmero y cúbito humano y dientes.Orgnac 3 (Francia). Decena de dientes aislados, parecidos a los de Arago. Aparece técnica de Levallois. 0,35-0,32 mda.Petralona 1. Cráneo casi completo. Apomorfias neandertales en órbitas, rostro hacia delante, frente y bóveda craneanas hacia atrás con una cara alargada y una mandíbula muy ancha. Descubierto por habitantes de Petralona (Grecia) en 1960. Su capacidad craneana es de 1220 c.c., alto para ser erectus, pero bajo para ser sapiens. Su edad se estima entre 0,25-0,15 mda. Hay quien considera que se trata del último Homo erectus, y otros le ven características de neandertal.Atapuerca. Sima de los Huesos – Ibeas de Juarros. Más de 5.000 fósiles (90% de los conocidos del Pleistoceno Medio en todo el mundo) de unos 28 individuos. 0,32 mda. Los primeros, descubiertos en 1976, por el equipo de T. Torres. En su mayor parte, corresponden a individuos jóvenes y adolescentes. El peso calculado por J. M. Carretero para jóvenes y adultos, está entre 58,75 y 81,43 kg. La estatura media de los varones, 1,74 m y la de las hembras 1,6 m. Para Emiliano Aguirre (2008) estos individuos se hallaban juntos en el interior de una cueva cuya entrada se cerró accidentalmente. Los restos fueron arrastrados por el fango y se deslizaron hasta la boca de la sima. Por el camino, algunos carnívoros penetraron por alguna rendija y dejaron sus huellas en los huesos. Esto explicaría también la ausencia de niños pequeños, cuyos restos habrían sido totalmente engullidos por los carroñeros. Los individuos, en general, estaban sanos aunque habían pasado por algunas crisis alimentarias; eran propensos a las afecciones artrósicas y a algo parecido a la osteoporosis; no hay rastro de accidente o enfermedad causante de la muerte.· 16 cráneos. 10 individuos padecían cribra orbitaria, extensión de perforaciones en el techo de la cavidad de las órbitas. En varios casos se observan traumatismos curados.· 20 mandíbulas (medio centenar de piezas), cuatro de ellas enteras. Son robustas, sin mentón. Cuerpo mandibular más bien bajo, con el borde inferior ancho, plano, paralelo a la fila de dientes y con un acusado reborde. Ramas ascendentes largas, poco altas y con el borde superior poco escotado. En seis individuos se observa una artrosis degenerativa de la articulación del hueso temporal y la mandíbula.· Más de 500 dienteso Incisivos superiores en pala.o Caninos con cínguos gruesos.o Premolares no molariformes.o Molares decrecientes, ya del primero al segundo, tanto en largo (igual que los neandertales) como en ancho (típicamente moderno).o Son frecuentes las hipoplasias en el esmalte. No se ven signos de caries, que aparecerían con el consumo de cereales, pero sí de la utilización de mondadientes. Hay marcas en la superficie externa de los dientes anteriores causadas por filos silíceos al utilizar la dentadura como herramienta.o Para María Martinón Torres y José María Bermúdez de Castro (2011) la morfología de estos dientes es completamente neandertal y por ello estos individuos de Atapuerca pertenecen a un grupo o especie diferenciados, pues estos rasgos están ausentes en los restos de los demás yacimientos de similar datación.· 42 restos de clavículas, 114 de escápulas, 645 restos de vértebras, 466 de costillas.o Las clavículas tienen como rasgo distintivo su longitud, su fuerte curvatura en ese y el fuste comprimido de modo anteroposterior, con sección ovoide.o Conducto medular largo en los atlas y corto en los axis, que son anchos en comparación con los modernos.· 110 restos de húmeros, 65 de cúbitos y 85 de radios.o En el húmero, la cabeza articular es oval con un ángulo de torsión bajo. Varios refuerzos en la zona articular del codo.· 173 restos de fémures, 20 de rótulas, 89 de tibias y 105 de peronés. Los huesos largos son muy fuertes.· 121 huesos del carpo, 103 del tarso, 93 metacarpianos, 80 metatarsianos, 208 falanges de pies y 304 falanges de manos. En los restos de niños, se dan varios casos de líneas de Harris, consecuencia típica de enfermedades infantiles.· Hioides y huesecillos del oído. I. Martínez ha deducido de su análisis la capacidad de habla de aquellos individuos.· Elvis. 0,5 mda. Pelvis muy ancha, con fuertes inserciones musculares correspondientes a una piernas muy robustas. La pelvis mejor conservada de todo el registro fósil, junto con otros restos del esqueleto, correspondiente a un varón muy corpulento que padeció importantes enfermedades degenerativas desde mucho antes de morir, con más de 45 años de edad. Dichas enfermedades debieron de producirle intensos dolores en las zonas lumbar y pélvica que le obligaban a caminar encorvado y, probablemente, a utilizar un bastón para mantenerse erguido. Debido a sus mermadas condiciones físicas, este individuo no estaba en condiciones de cazar ni de llevar a cabo otras tareas útiles para su comunidad, lo cual ha llevado a la conclusión de que, además de alimentación, el anciano debió también de recibir ayuda para seguir el ritmo de la marcha de su gente, un grupo de cazadores nómadas que se desplazaban junto a las manadas de herbívoros de los que se alimentaban. Un tipo de conducta social de ayuda a los más desvalidos de la que existen muy pocos indicios para épocas tan remotas pero sobre la que en Atapuerca ya se ha encontrado más de una prueba.o En efecto, en marzo de 2009 se publicó un estudio que demostraba que un niño con el cráneo deforme, por fusión prematura de los huesos del cráneo, de unos 10 años (que los científicos bautizaron como Benjamín) y que pertenecía a la misma especie y época del anciano, fue cuidado y mantenido con vida todo el tiempo posible por sus congéneres.· Miguelón, Atapuerca 5 (AT 700). Descubierto en la Sima de los Huesos (Atapuerca) entre 1992 y 1993 por Juan Luis Arsuaga. Su antigüedad se estima en 0,3 Ma; su capacidad craneal es de 1.125 c.c. Se trata del cráneo más completo de heilderbengensis descubierto hasta ahora. (Arsuaga et al. 1993; Johanson and Edgar 1996). Se ha identificado la mandíbula que le corresponde.o Macizo maxilar grande, proyectado hacia delante sin fosa canina entre el saliente nasal y el pómulo ni fosa maxilomalar entre el pómulo y la pared de las raíces de los molares superiores. Orbitas grandes y casi redondas. Prognatismo y adelantamiento del arco de los dientes que deja un espacio retromolar entre el talón de la última muela inferior y el borde anterior de la rama ascendente de la mandíbula.o Padecía varias alteraciones en la boca. La más grave, una infección en el lado izquierdo tras la fractura de un diente.o Del estudio de la lateralidad del cerebro, se concluye que era diestro.· Atapuerca 4. Caja craneal completa, sin su parte facial. Se observa una artritis avanzada en los cónditos occipitales, con importante dificultad y dolor en los movimientos de la cabeza y los dos conductos auditivos obstruidos por hiperostosis.· El frontal huido, la bóveda baja y el occipital alargado y anguloso hacia atrás del cráneo 5 no son compartidos en el cráneo 4. La bóveda relativamente baja y el frontal estrecho son rasgos comunes.Atapuerca-Galería, en la comunicación con la Sala de los Zarpazos. 0,3 Ma.· AT 76-T1H. Fragmento de mandíbula hallado por Trinidad Torres (1976). La presencia de espacio retromolar, el toro mandibular y los tamaños de los últimos molares jutifican su asignación al demo de los preneandertales europeos.· Fragmento de cráneo hallado en 1995 que comprende la parte posteromesial del parietal derecho, parte posterior de la sutura sagital, otro fragmento menor del parietal izquierdo y el tramo central de la sutura labdática, con una porción muy pequeña adherida del occipital que incluye el lambda. El parietal es muy grueso y fue hundido por un golpe que luego sanó.Boxgrove (Reino Unido). Excavado por Mark Roberts en los 90. El yacimiento se mantuvo intacto desde que hace 0,5 Ma fuese cubierto de forma rápida por el fango, que permitió su conservación. Se han encontrado huesos de elefante, rinoceronte, caballo, ciervo, león, lobo, hiena, castor, liebre, ratón de campo y rana. Muchos huesos conservan huellas líticas y se ha podido reconstruir el proceso de matanza o despiece. Para Roberts, los animales fueron cazados mientras que Gamble defiende que son fruto de la carroña. Los únicos restos humanos son dos dientes y parte de una tibia (0,35-0,3 Ma). No hay hogares ni señales de estructuras.Castel di Guido (Italia). Fragmentos craneales. Una diáfasis de fémur. Asociados a industria modo 2. 0,3 Ma.Ponte Mammolo (Italia). Un fémur. 0,2 Ma.Pech-de-l’Aze (Francia). Infantil. 0,4-0,2 Ma.Biache-Saint Vaast (Francia) Cráneo incompleto. Trazado de la red meníngea avanzado, parecido con Swanscombe y con neandertal. 0,263-0,175 mda.Swanscombe (Reino Unido). Parietal y occipital de hembra joven adulta. 1.300 cc. Formas muy próximas a neandertal. 0,25 mda.Pontnewydd (Reino Unido) Fragmentos de maxilar y tibia. Dientes. 0,251-0,195 mda.Steinheim (Alemania) Cráneo casi completo, deformado y restaurado. Apomorfias de neandertal poco avanzadas. 0,225 mda.Ehringsdorf (Alemania) Restos craneales y poscraneales de varios individuos. Rasgos de neandertal avanzados. 0,25-0,2 mda.Grotta del Principe (Italia). Pelvis de individuo femenino. 0,2 mda.Sedia del Diavolo (Italia). Fragmentos de cráneo y fémur y un metatarsiano. 0,2 mda.Grotta del Poggio (Italia). Un diente y varios poscraneales. 0,2 mda.Casal de Pazzi. Fragmento de parietal. 0,2 mda.Le Lazaret (francia). Paietal de inmaduro, con cicatriz de trauma y meningioma postraumático; un incisivo lacteal y un canino adulto. Asociado a industria modo 2 evolucionado. 0,16 mda.Apidima I (Grecia). Dos cráneos. 0,2 mda.Apidima II (Grecia). Rostro, frente y bóveda craneal muy próximos a neandertales de Saccopastore y Gibraltar.Grotte Suard. La Chaise (Francia). Pates de un cráneo adulto, otro infantil y su mandíbula, tres dientes. Asociados a industria tipo 2 evolucionado, con lascas Levallois. 0,13 mda.Cueva del Congosto (Alcorlo, Guadalajara, España). Un metatarsiano y una vértebra cervical. Asociados a restos de animales con trazas de haber sido un basurero de hienas.Cueva de Lezetxiki (Mondragón, Guipúzcoa, España). Húmero completo de un individuo femenino. Datación incierta.Reilingen (Alemania). Cráneo incompleto. Algunos rasgos afines a neandertal.Montmaurin (Francia). Mandíbula. Rasgos de preneandertal.Características.
- Dientes con tamaño y corona reducida, mucho más pequeños que el erectus.
- Caja craneal voluminosa y en pendiente. Frente deprimida, con toro poderoso en dos partes. Bóveda craneal bastante baja y alargada. Occipucio anguloso con repliegue.
- Prognatismo profundo. Arco cigomático saliente y orientado hacia delante.
- Arcada dental parabólica. Mandíbula moderadamente robusta.
- Reducción del tamaño de los molares.
- Pelvis corta y en forma de cubeta. Miembros inferiores relativamente largos.
Inferencias sobre su modo de vida.
Más sobre Homo heidelbergensis y Homo neanderthalensis.
- Regiones templadas o cálidas. Paisajes variados de árboles o agua, pero no jungla densa.
- Modo de vida y dieta: cazadores capaces de matar animales grandes mediante emboscada; recolectores de pequeños animales, huevos y todo tipo de plantas silvestres comestibles. De su higiene sabemos que usaban palillos de dientes para la limpieza de los espacios interdentales. Usaban abrigos de piel para evitar el frío y fabricaban herramientas de piedra y madera. En Schöningen se hallaron lanzas de madera datadas en 0,4 Ma.
- Campamentos: construcción de cabañas de madera y quizás pieles de animales, pero también uso de abrigos y vestíbulos de cuevas.
- Organización social: pequeños grupos o clanes de entre 15 y 40 individuos jerarquizados, que explotaban y defendían los recursos de un territorio y que realizaban contactos esporádicos con otros grupos para evitar la endogamia. Probablemente podían comunicarse mediante el habla.
Desnudos por la Educación.
 |
| Movilización lationamericana por la Educación. Cartel. |
 |
| Movilización latinoamericana por la Educación. Concepción. |
Jóvenes estudiantes se concentraron en Concepción el 24 de noviembre de 2011, para pedir una Educación pública, en el marco de una movilización latinoamericana. En sus cuerpos, las banderas de cada unos de los países participantes de la movilización.
 |
| Movilización latinoamericana por la Educación. Concepción. |
http://www.lanacion.cl/jovenes-protestaron-desnudos-en-el-centro-de-concepcion/noticias/2011-11-24/173811.html
http://www.biobiochile.cl/2011/11/24/con-sus-cuerpos-desnudos-pintados-con-banderas-americanas-jovenes-protestan-por-la-educacion.shtml
viernes, 25 de noviembre de 2011
Later Stone Age y aparición del comportamiento humano moderno.
Henshilwood y Marean (2003) sintetizaron los rasgos de la conducta humana moderna:
Enterramientos y prácticas rituales.
Arte, adornos y decoración.
Uso simbólico del ocre.
Trabajo del hueso y el asta.
Tecnología lítica laminar.
Estandarización de los tipos de artefactos.
Diversidad de artefactos.
Construcción de hogares estructurados.
Uso organizado del espacio doméstico.
Redes amplias de intercambio.
Explotación efectiva de grandes mamíferos.
Movilidad estacional.
Ocupación de ecosistemas rigurosos.
Pesca y caza con proyectiles.
Los yacimientos africanos son más abundantes entre 80.000-60.000 años, que en el periodo posterior, lo que dificulta las investigaciones. Deacon y Wurz (2001) señalan que el último interglacial (5e) fue particularmente benigno en África, lo que facilitó la extensión de la MSA por todo el continente. Entre el 60.000-30.000 la mayoría de los yacimientos sudafricanos fueron abandonados debido a la aridez asociada a la última glaciación. El patrón europeo es el contrario, lo que seguramente está relacionado con una migración importante de grupos modernos desde África.
Teorías sobre la aparición del comportamiento moderno.
Teoría tradicional.
El comportamiento humano moderno aparece hace 50.000-40.000 años, cuando en el registro arqueológico la Middle Stone Age es sustituida por la Later Stone Age.
Se produjo una sustitución biológica de poblaciones y la desaparición de los neandertales.
La conducta moderna apareció repentinamente (cambio puntuado) quizá debido a una mutación de efectos neurológicos.
La MSA se identifica con el Paleolítico Medio de los sapiens arcaicos europeos y la LSA con el Paleolítico Superior.
Está apoyada por Bar-Yosef, Davidson, Noble, Stringer, Gamble, Mellars, etc.
Para Klein (2001) los yacimientos típicos de la MSA de finales del Pleistoceno Medio y comienzo del Superior (Blombos Cave, Klaises River, Border Cave…) se caracterizan por lascas más o menos alargadas extraídas de núcleos preparados; entre los retocados predominan raederas, denticulados, puntas bifaciales y elementos de dorso. Estos hallazgos son muy semejantes a los del musteriense y Klein no acepta las tradiciones regionales. MSA se diferencia de LSA claramente en la menor variedad de tipos líticos, estabilidad y uniformidad de los conjuntos en el tiempo y el espacio, ausencia de objetos de hueso, concha y manifestaciones artísticas. Los artefactos en hueso y conchas perforadas con restos de ocre hallados en Blombos Cave y atribuidos a la MSA (70.000 años) podrían deberse a una mezcla de los materiales estratigráficos. Durante la LSA hay una intensa actividad pesquera y de caza de pájaros, lo que manifiesta una tecnología sofisticada, que no existe de ningún modo durante la MSA. Los restos de conchas son mucho más grandes en la MSA lo que indica una recolección mucho más intensiva durante la LSA. Durante la LSA, los cazadores recolectores cazaba focas en la costa entre agosto y octubre, cuando las crías de 9-10 meses pueden ser interceptadas junto a la línea del mar. Por el contrario, en la MSA la movilidad estacional parece no existir, lo que Klein interpreta como una dependencia de las fuentes de agua. Por último, en la LSA los humanos cazaban animales peligrosos gracias al uso del arco y la flecha.
Últimas teorías.
Los nuevos modelos sugieren que la conducta moderna cambió de forma gradual, al mismo tiempo que el morfotipo del sapiens. Sí aceptan una sustitución abrupta del Paleolítico Medio por el Paleolítico Superior en Europa pero no de la MSA por la LSA.
El modelo gradual, sostiene que la capacidad de apreciar las formas bellas achelenses se va desarrollando de manera gradual y continua. El origen del arte es difuso, extendido en el espacio y continuo en el tiempo.
La mayor crítica es la escasez de restos artísticos en el Paleolítico Medio que contrasta con la explosión en el Paleolítico Superior.
McBrearty y Brooks (2000)
La naturaleza revolucionaria del Paleolítico Superior europeo se debe más a la discontinuidad del registro arqueológico que a las transformaciones culturales y cognitivas. Son los yacimientos africanos más antiguos los que deben darnos el punto de partida Entienden que los sapiens arcaicos engloban con probabilidad varias especies.
En la MSA existe un fuerte componente laminar en conjuntos muy antiguos como Gademotaa o Kapthurin.
Reconocen las tradiciones culturales.
Destacan las industrias geométricas y microlíticas en contextos de MSA.
Aceptan que los artefactos en hueso hallados en Blombos Cave pertenecen a la MSA.
Interpretan que la mayor abundancia de caza de animales peligrosos en la LSA se debe a una ampliación del espectro dietético debida a la presión demográfica. De modo parecido opinan Henshilwoodd y Marean (2003).
Para Cela y Ayala, este es el esquema más razonable para entender las evidencias arqueológicas, paleontológicas y moleculares. La nueva mente y quizá el lenguaje son patrimonio de nuestra especie a lo largo de todo el lapso de su existencia.
Las pruebas que pueden presentarse son:
o Enterramientos
con cáscaras de avestruz y huesos perforados (collares) de hace 100.000 años en
Border Cave (Sudáfrica) y Zhoura (Marruecos).
o Piedras
ornamentales en Seggédim (Níger 130.000 años).
o Muchos
yacimientos con cuentas, objetos grabados y perforados (40.000-20.000 años).
o Para
Ambrose y Lorenz (1990) las puntas de tipo Still Bay y los conjuntos Howiesons
Poort (70-60 ka) representan una muestra de la capcidad de los humanos modernos
para responder a los cambios en la distribución de los recursos producidos por
las transformaciones de las condiciones ambientales.
o Serie
de arpones en hueso del yacimiento de Katanda (Congo) y artefactos de madera de
Blombos Cave (70 ka).
o
Explotación de recursos marinos y fluviales.
Se han recuperado restos de peces, moluscos o artefactos de pesca en Blombos,
Katanda, Klasies River, Die Kelders y Sea Harvest. En 2011 se publicó el
hallazgo en la cueva de Jerimalai, en la isla de Timor Oriental, de 38.000
huesos de 2.843 peces de 23 especies diferentes capturados por humanos hace
42.000 años. Entre los restos hay huesos de atunes, tiburones y rayas, lo que
significa que estos primeros humanos ya pescaban mar adentro y en las
profundidades.
o Hay
pruebas de estructuració del espacio doméstico en Katanda, Mumbwa Cave (Zambia)
Rose Cottaghe Cave (Sudáfrica), Rhafas (Norte de África) y Wadi Sodmein (Norte
de África).
o Pigmentos
ocres y rojizos metálicos.
ü Podrían
haberse utilizado para decorar el cuerpo humano. Knight y colaboradores (1995)
han sugerido que la decoración del cuerpo de los machos podría ser una
representación simbólica de la menstruación femenina y tener así funciones de
organización social coincidentes con el inicio de la división del trabajo.
- Estas evidencias indican tres cosas:
o Los cambios cognitivos de Homo sapiens se traducen en indicios de una cultura moderna desde la aparición de nuestra especie en África.
o El aumento de complejidad fue gradual.
o Sólo dejando de lado los indicios africanos es posible asombrarse ante la aparición súbita de la explosión artística auriñaciense.
- Las tesis gradualistas hablan de manifestaciones artísticas o simbólicas ya en el Pleistoceno Medio, con los ergaster. McBrearty y Brooks (2000) reducen al sapiens la conducta simbólica.
- Los gradualistas tendrán que aceptar que las manifestaciones supuestamente artísticas de los periodos achelense (grado erectus) y musteriense (Neandertal) son muy escasas de significado dudoso y, por así decirlo, cogidas por los pelos en cuanto a su contenido simbólico. En realidad lo que hacen Sally McBerry y Alison Brooks es ampliar la explosión de la eclosión artística de 40.000 a 100.000 años pero siempre dentro de la autoría por Homo sapiens. Algo que daría la razón a estas antropólogas sería la recién descubierta Venus de Tan Tan en Marruecos una figurilla vagamente antropomorfa de mas de 300.000 años de antigüedad pero parece ser una obra del azar por erosiones después modificada con alguna incisión por el Homo al encontrar esta figura de 5 cm. de altura con alguna forma humana. El problema es que este hallazgo como otros similares es aislado y excepcional mientras que la “explosión” artística del Paleolítico Superior es extensa y masiva.
miércoles, 23 de noviembre de 2011
Zuloaga. Desnudo de récord.
 |
| Ignacio Zuloaga. Madame Souty reclinada en un sofá. |
El cuadro de Ignacio Zuloaga Madame Souty reclinada en un sofá fue adjudicado en una subasta en Sotheby's (Londres) el 22 de noviembre por 829.368 €, un precio récord para una obra del artista español.
El cuadro es un retrato de la modelo favorita de Zuloaga en su estudio de París, pintado en 1921 y pertenecía a la colección privada del autor.
martes, 22 de noviembre de 2011
Ocularpation. Desnudando la avaricia de Wall Street.
 |
| Ocularpation. Foto de Hal Horowitz. |
 |
| Ocularpation. Foto de Hal Horowitz. |
Ocularpation es una inciativa de Zefrey Throwell, desarrollada con actores, para denunciar la avaricia de Nueva York. Tuvo lugar el 1 de agosto de 2011.
 |
| Ocularpation. Foto de Anthony Miller. |
 |
| Ocularpation. Foto de Anthony Miller. |
http://www.artnet.com/magazineus/news/artnetnews/zefrey-throwell-ocularpation-wall-street-8-3-11_detail.asp?picnum=14
http://artistsspeakout.com/2011/08/50-performance-artists-get-naked-three-get-arrested-protesting-corruption-on-wall-street/
El 18 de noviembre, Zefrey Throwell, organizó un strip poker para apoyar la ocupación de Wall Street.
Strip Poker in Manhattan - I'll Raise You Two! por ramimor
Evolución del cerebro en los homínidos.
Los estudios
alométricos se centran en:
-
La
relación entre el tamaño cerebral en peso y la superficie corporal en área.
-
La
relación entre la extensión del neocórtex y el volumen de la médula espinal
(Passihgham, 1975).
El incremento de la
capacidad craneal puede ser un buen indicio acerca de la aparición de capacidades
cognitivas nuevas, sobre todo si tenemos en cuenta lo caro que es, en términos
metabólicos, un órgano como el cerebro.
Se apunta de manera
muy generalizada a una expansión del córtex frontal (Deacon, 1997). Sin embargo,
Semendeferi y Hanna Damasio (2000) sometieron a prueba la hipótesis mediante la
comparación en distintos hominoideos de los volúmenes relativos de tres áreas cerebrales,
la frontal, la temporal y la parieto-occipital (definidas por el sulcus central
y la fisura de Silvio) para concluir que en ninguna de las tres existe una expansión
extraalométrica en nuestra especie. Tenemos el área frontal que corresponde al tamaño
de nuestro cerebro. Por fortuna Rilling e Insel (1999) han rescatado en alguna forma
la idea correcta gracias a su estudio
acerca de la girificación. La girificación es la relación que existe, en cualquier
sección del córtex, entre la longitud del perímetro cortical exterior (trazando
el contorno más externo del córtex sin seguir sus recovecos) y la del perímetro
total (midiendo el interior también de los plegamientos que hacen que el córtex
humano parezca algo similar a una coliflor, salvadas sean todas las distancias).
Rilling e Insel descubrieron que la girificación es, en términos generales, alométrica
en los hominoideos: cuanto más grande es el cerebro, más girificado está el córtex.
Pero en las secciones longitudinales horizontales Rilling e Insel detectaron una
girificación extraalométrica en la “rebanada” que corresponde en gran parte al córtex
frontal y prefrontal en Homo sapiens. Eso indica que el desarrollo último del lenguaje,
la capacidad estética y los juicios morales puede tener que ver con esas áreas,
aunque están por realizar los experimentos de localización que sean capaces de sacarnos
de las dudas. Entretanto, Zeki (1999) sugiere que el arte pictórico podría deberse
a ciertos cambios producidos en las mismas áreas visuales, es decir, en la zona
occipital del cerebro.
No existe ningún lazo directo entre tamaño cerebral
y habilidades cognitivas y los paleontólogos han intentado encontrar, sin éxito
completo, unos parámetros que pudieran expresar la inteligencia. Los antropólogos se refieren a tamaños craneales de
las distintas especies y suelen acudir a procedimientos estadísticos.
-
El coeficiente de encefalización mide la relación
existente entre el peso del cerebro y el peso del cuerpo). No es disparatado
atribuir tanto un cerebro mayor como unas capacidades cognitivas más complejas
a la especie que tiene una encefalización más alta.
o
Entre los monos del Nuevo Mundo, los que tienen
alimentación insectívora cuentan con cerebros más grandes que los que se
alimentan de hojas y ese hecho se relaciona con la mayor necesidad de
procesamiento de la información procedente del medio ambiente.
o
Una suposición paralela es la que lleva a atribuir
al habilis la manufactura de las
primeras herramientas porque su coeficiente de encefalización es superior al
del Australopithecus boisei.
-
Los primeros homínidos tienen unas capacidades
craneales semejantes a las de los chimpancés actuales. Éstos son capaces de
utilizar herramientas simples como palos o piedras e incluso establecen
distintas tradiciones de manipulación aprendida de objetos, pero no parecen
disponer de la capacidad de anticipación, planificación y coordinación
necesarias para llevar a cabo la cultura olduvaiense.
Encefalización en Homo.
La espina dorsal rudimentaria
de los primeros vertebrados se dividió en cinco partes, medula, cerebro posterior,
cerebro medio, diencéfalo (tálamo e hipotálamo) y telencéfalo (hemisferio cerebral).
A lo largo de la filogénesis posterior tan solo cambiaron los desarrollos relativos
de esas regiones, sobre todo con la expansión de los centros olfatorios del telencéfalo
que darán lugar, en los mamiferos, a la aparición del neocórtex.
Los mamíferos más primitivos
se distinguen de los reptiles por la posesión de un córtex bien desarrollado, con
diversas capas de neuronas, en el que Elliot Smith identificó tres regiones, a
principios del XX. Dos de ellas podían identificarse con regiones semejantes de
los vertebrados inferiores, pero la otra es un rasgo único de los mamíferos,
con filogénesis en forma de mosaico: el neocórtex, con cambios evolutivos
dispares en los distintos linajes.
Una característica general
de todos los primates es el gran desarrollo alcanzado por el neocórtex y dentro
de él de las áreas visuales (expansiones extra alométricas, superiores al
aumento de tamaño del cuerpo).
Australopithecus
africanus disponía de una capacidad encefálica en torno a
los 450 cc. Homo habilis, con un
tamaño corporal similar al A. Africanus ,
600. El cráneo KNM-ER 1470 tiene una capacidad de 752 cc., está datado hace
1,9 Ma y atribuido a Homo rudolfensis. Para
José María Bermúdez de Castro, (La
evolución del talento) el incremento en el coeficiente de encefalización
pudo producirse:
·
Gracias a un aumento en la en la tasa de
crecimiento del cerebro durante la gestación. La pelvis del Homo habilis era relativamente más ancha
que la del sapiens, y el aumento del
tamaño del cerebro no debió suponer problema alguno en el parto.
o
La morfología de los huesos de la pelvis, el
insquion y el pubis, indican que en el Australopithecus
afarensis el parto era parecido al de las humanas modernas, con rotación de
la cabeza del feto y una trayectoria curva del canal del parto. El feto tenía
un 50% de posibilidades de nacer con la coronilla orientada hacia su madre, lo
qu dificulta el parto en solitario.
o
Los problemas crecieron cuando los adultos
alcanzaron un tamaño cerebral de 800 cc., con Homo erectus. Debido a la morfología del útero, la vagina y la
pelvis, el parto tuvo que producirse casi siempre con la coronilla del feto
orientada hacia la madre. El canal del parto tiene un diámetro máximo de 13 cm.
La asistencia al parto es casi obligada. A partir de entonces los incrementos
en el tamaño del cerebro tuvieron que producirse fuera del útero de la madre y
esto pudo llevar a un aumento de la altricialidad o inmadurez de la cría en el
momento del parto.
·
Gracias al aumento en la duración de la infancia.
El crecimiento del cerebro se prolonga cierto tiempo después del nacimiento. En
los chimpancés, el cerebro crece 250 cc., durante su infancia, de unos cinco
años. La prolongación de la infancia implica la prolongación de la lactancia y
el aumento del intervalo entre nacimientos con la consiguiente disminución de
la fertilidad.
o
La solución evolutiva a este compromiso entre el
aumento del tiempo de infancia y el mantenimiento de la fertilidad fue la
progresiva aparición de la niñez, cuando la cría no precisa de lactancia
mientras mantiene el crecimiento.
§
Homo habilis y rudolfensis dispondrían de una infancia
de hasta cinco años y un periodo juvenil de otros cinco años que daría paso a
la aptitud para la reproducción sexual.
·
El periodo de infancia pudo reducirse gracias a la
incorporación de las grasas animales a las dietas de las crías. Esta dieta, más
rica, pudo abastecer los requerimientos energéticos de un cerebro más grande.
§
Según Trinkaus, el desarrollo dental de un niño
neandertal de 8 años era comparable al de un niño sapiens de varios años más.
§
Los humanos actuales disponemos de un tiempo de
desarrollo de alrededor de 18 años que incluye dos etapas nuevas: niñez y
adolescencia.
·
Nueve meses de desarrollo fetal.
·
Infancia: lactancia y dentición decidua.
Crecimiento muy rápido del cerebro. El primer año, a una velocidad similar a la
que se produce durante el desarrollo fetal.
·
Niñez o segunda infancia. Hasta los seis o siete
años, con la aparición de los primeros molares defintivos. A los siete años, el
cerebro alcanza el tamaño defintivo.
·
Juventud. Hasta los once o doce años, cuando surge
el segundo molar permamente
·
Adolescencia.
·
Es posible que los genes que participaron en estos
cambios fuesen el MCPH1, el ASPM y el NRCAM
Encefalización y lenguaje.
El paso de hominoideos primitivos a seres del género
Homo tiene que ver con el desarrollo de
habilidades de modificación del entorno: construcción de herramientas, dominio del
fuego, cooperación y división del trabajo etc. Pero estas actividades necesitan
de un lenguaje con el que la transmisión de experiencias de haga al margen de la
herencia genética a través de una memética cultural.
Los sapiens cuentan con un desarrollo muy
acusado de las áreas temporales y prefrontales relacionadas con el
procesamiento de la comunicación verbal y las tareas semánticas. Cabría
sustentar en el desarrollo de tales áreas las especulaciones acerca del origen
del lenguaje.
-
Tobias
ha venido defendiendo una expansión temprana de estas áreas. Comparando los
endocráneos de Homo habilis con los
de Australopithecus africanus, Tobias
detectó no solo un desarrollo de las áreas de Broca y Wernicke sino también del
lóbulo frontal, que anticiparía el incremento de volumen de esa zona en los
homínidos de grado erectus.
o
Más
allá de los indicios extraídos de los moldes endocraneales, apenas puede
decirse nada que no sea en términos especulativos. Lieberman sostiene que el habilis tendría, en el mejor de los
casos, un lenguaje no totalmente moderno mientras que Tobias cree en una
creciente complejidad en los aspectos conceptual y sintáctico, junto con una
ampliación en el tiempo del repertorio de fonemas. Todas estas etapas entrarían
dentro de lo que cabe considerar como “lengua humana hablada”.
-
Holloway,
Jerrison, Falk y Tobias indican la presencia de dos fronteras en la evolución de
la complejidad cerebral.
o
La
primera, la aparición en Homo habilis
de una organización neurológica “esencialmente humana”.
o
La segunda,
un continuo y rápido aumento del índice de encefalización dentro del género Homo.
-
Aiello
y Dunbar (1993) buscan una evidencia medible, como el tamaño cerebral, para
rastrear el origen del lenguaje. La conclusión apunta a las primeras fases del
género Homo como protagonistas de la
aparición del leguaje, con un rápido incremento en la segunda mitad del
Pleistoceno Medio, dentro del grado erectus.
Establecieron una relación entre el tamaño de la corteza cerebral con
respecto al resto del cerebro, el número de individuos que forman los grupos y
el tiempo dedicado a la interacción social. En grupos muy numerosos (se supone
que los grupos de habilis pudieron
ser de más de 100 individuos) la jerarquía y el liderazgo ayudan a conservar la
unidad; entonces resulta indispensable cierta complejidad en la comunicación.
o
Robert
Martin (2000) supone una escasa diferenciación sexual en el Australopithecus africanus y por ello
cree que se sobreestimó su tamaño corporal, de modo que el incremento de
encefalización se daría ya en esta especie, posiblemente relacionado con la
actividad locomotora y la búsqueda de un tipo diferente de alimentos a su vez
relacionados con las necesidades de un cerebro en expansión.
o
Frank y
colaboradores (2000) comparando los endocráneos de diferentes especies de
parántropos y los de Austraopithecus
africanus dan una idea de la evolución cerebral parecida a la de Martin,
apoyando la hipótesis del Australopithecus
africanus como un ancestro del Homo
-
Para
Semendeferi, Damasio y Frank, tanto el tamaño del lóbulo frontal como la
distribución de sus sectores es muy parecida en los monos, en los simios y en
los seres humanos. La evolución del linaje humano implicó un aumento en el tamaño
cerebral, pero no un desarrollo relativo del lóbulo frontal que se supone interviene
en algunos procesos importantes para el lenguaje como son el pensamiento creativo,
la planificación de acciones futuras, la expresión artística, o el análisis semántico.
Nuestra área frontal es, en términos relativos, la que corresponde a un primate
con un cerebro de nuestro tamaño y solo el gibón tiene un área frontal menor en
términos alométricos. La conclusión es que las modificaciones evolutivas del
córtex frontal son anteriores a la separación chimpancés-humanos, así que las
diferencias cognitivas entre simios y humanos no pueden estudiarse de esta
manera.
o
El
estudio de Rilling e Insel (1999) indica en contra de lo anterior, que se ha
producido una expansión extraalométrica de la girificación (plegamientos) en
dos zonas corticales del cerebro humano: la temporal parietal/posterior y la
prefrontal.
o
El
estudio comparativo de Falk y colaboradores (2000) indica una medida relativa
igual para el lóbulo central en humanos actuales, gorilas y chimpancés, pero
con diferencias significativas a favor de los humanos en algunas subregiones
del lóbulo central. Esto quiere decir que el lóbulo frontal humano se ha
reorganizado en comparación con el de los grandes simios africanos. Los lóbulos
frontales de los parántropos muestran una forma tipo-simio mientras que la
expansión frontal y temporal de Australopithecus
africanus se acerca más a Homo.
Será necesario
disponer de estudios más detallados para localizar de forma precisa las
funciones cerebrales que intervienen en los procesos cognitivos relacionados
con el habla antes de poder decir la última palabra.
LATERALIZACIÓN.
El hecho de la encefalización creciente no es el único fenómeno
notorio que pueda detectarse en la filogénesis de los homínidos. Además del aumento
de tamaño relativo, que es una cuestión en esencia cuantitativa, se producen también
cambios en la organización que podriamos llamar cualitativos siendo el más importante
la lateralización.
La lateralización supone
una quiebra de la asimetría básica de los dos hemisferios cerebrales. Así los centros
de control del lenguaje está lateralizado: excepto en el caso de algunos
zurdos, se encuentran en el hemisferio cerebral izquierdo. Si damos por bueno
el modelo de Finlay y Darlington (1995) acerca de la evolución conjunta de
muchos de los rasgos distintivos de la conducta humana bajo control del
cerebro, podemos suponer que esa lateralización, relacionada hoy con el
lenguaje, pudo aparecer por otros motivos. Cualquier dato acerca de la
aparición de la tendencia a utilizar la mano derecha se convierte en una evidencia
acerca de la lateralización en su conjunto.
-
Mediante
el estudio de las marcas dejadas en las esquirlas obtenidas al fabricar las
herramientas líticas, Toth (1985) concluyó que la tendencia a utilizar la mano
derecha está presente en la cultura olduvaiense hace 2 mda. Toth y Schick
(1993) sugieren una relación con el desarrollo de las áreas lingüísticas.
-
El
sentido de los modelos que ligan lenguaje, destreza y lateralización cerebral
puede someterse a prueba mediante el examen de otros primates. Se ha seguido
tradicionalmente la tesis de Warren (1980) de que la lateralización es un
fenómeno que solo aparece en nuestra especie.
o
Los
chimpancés no muestran preferencia por el uso de una mano en particular. Sin
embargo, puede constatarse un predominio del planum temporale del hemisferio izquierdo, un componente esencial
del área de Wernicke de los humanos (Gannon, 1998). Partiendo de la hipótesis
de una asimetría fijada por los antecesores comunes de humanos y chimpancés se
pueden seguir cuatro diferentes líneas de interpretación respecto a la
intervención de la lateralización en el desarrollo del lenguaje.
1.
Sostener
que esta asimetría de los antecesores no tuvo ninguna función comunicativa.
Esta función aparecería más tarde solo en los humanos. La función que tiene en
los chimpancés es desconocida.
2.
La
asimetría tuvo funciones de comunicación. En los chimpancés permite una
comunicación visual basada en gestos.
3.
El planum temporale izquierdo carece de
función comunicativa, antigua o moderna.
4.
Se
trata de una homoplasia. Esta es la menos parsimoniosa de las interpretaciones.
o
Merchant
y McGrew (1998) señalan que los chimpancés cautivos parecen comportarse como si
existiera preferencia por la diestra mientras que los que permanecen en su
hábitat original solo muestran esta preferencia en el manejo de herramientas.
Indican que una parte de nuestras convicciones acerca de la lateralización y su
filogénesis puede basarse en supuestos que no están bastante contrastados.
o
Peña-Melián,
Antonio Rosas et al analizaron tres
cráneos fósiles de Homo neanderthalensis
encontrados en el yacimiento de El Sidrón (Asturias) (2011) encontrando que el
cerebro de los neandertales era asimétrico, más que el del ser humano actual.
Etapas en la evolución del cerebro.
La evolución del cerebro es vista por Holloway a través de diversas etapas:
- Reorganización del parietal posterior y del occipital anterior
hacia pautas más humanas, pero sin cambios apreciables por lo que hace al
volumen cerebral, que continúa en Australopithecus
dentro de los valores de los simios.
- Hacia los 2 Ma el linaje Homo,
cuenta con un tercio inferior del frontal más complejo y moderno en
sus circunvoluciones, un volumen craneal superior y asimetrías cerebrales
similares a los de los seres humanos actuales.
- Todos los primates antropoides presentan asimetrías morfológicas
entre los hemisferios cerebrales.
- En el humano moderno, estas asimetrías se corresponden con
especializaciones en el lenguaje simbólico (hemisferio izquierdo) y la
integración visuo-espacial (derecho), lo que hace plantear a Holloway la
posibilidad de que tales capacidades existiesen ya en los primeros Homo, aun cuando menos
desarrolladas. Esta es una idea expresada por primera vez por Tobias.
- La asimetría cerebral tiene también una expresión en términos de
dimorfismos sexuales. La relación estrecha madre-hijo en los homínidos,
incluso acentuada por una posible prolongación del periodo del embarazo,
llevaría a una especialización cerebral relacionada con el corpus callosum. Funcionalmente
hablando, las diferencias tenderían a establecer un cerebro masculino
especializado en la integración visuo-espacial relacionada con la caza.
Por el contrario, las hembras tendrían unas superiores capacidades
cognitivas respecto a la comunicación simbólica y la comprensión de la realidad
social.
i. Estas habilidades
serían complementarias más que competitivas.
ii. La manera como los
dimorfismos cognitivos podrían haber afectado a la evolución humana es
imposible de plantear como no sea en términos especulativos.
Reducción reciente del tamaño del cerebro.
Hace 40 ka, el cerebro del sapiens ocupaba 1.500 cc. Desde entonces se ha ido reduciendo hasta
los 1.350 cc que tenemos hoy en día.
Son varias las hipótesis para explicar esta reducción:
- A medida que las sociedades se vuelven más complejas el tamaño del cerebro se reduce, porque aquellos individuos de la especie, que antes no hubieran podido sobrevivir por cuenta propia, ahora logran hacerlo con la ayuda de los otros.
- La reducción puede estar relacionada con la mayor eficiencia de nuestro cerebro.
- Los animales domesticados tienen cabezas más pequeñas que las de sus ancestros salvajes.
Suscribirse a:
Comentarios (Atom)













{kind=link}
{kind=link}
{kind=link}